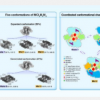
哺乳动物线粒体电子传递链复合物是线粒体中最重要和最复杂的蛋白质机制。尽管这个系统已经被研究了一个多世纪,但其组成和分子机制仍然很大程度上未知。在这里,杨的研究小组报告了猪呼吸链巨型复合物-I 2 III 2 IV 2 (MCI 2 III 2 IV 2 )五种不同构象的高分辨率冷冻电镜(cryo-EM)结构,包括状态1、状态2 、中 1、中 2 和中 3。高分辨率冷冻电镜成像与超分辨率门控受激发射损耗显微镜 (gSTED) 相结合,有力地支持了活细胞中MCI 2 III 2 IV 2的形成。每个MCI 2 III 2 IV 2结构包含141个亚基(70种不同的肽,2.9 MDa),总共240个跨膜螺旋。 MCI 2 III 2 IV 2结构中显示的 CI、CIII 和 CIV 之间的相互影响表明该巨型复合物可以作为电子转移和质子泵浦的整体单元。不同状态的构象变化表明了 MCI 2 III 2 IV 2激活/失活过程的合理调节机制。

杨教授课题组解析了猪MCI 2 III 2 IV 2 5种构象的高分辨率结构,即State 1、State 2、Mid 1、Mid 2和Mid 3,根据CI的构象、CIII的Rieske结构域进行分类,以及 CIV 的立场。根据CIV的位置, MCI 2 III 2 IV 2分为两种构象:扩张构象和收缩构象。在 MCI 2 III 2 IV 2的收缩形式中发现的两个 CI均处于失活状态。相比之下,MCI 2 III 2 IV 2扩展构象中的两个 CI可以处于活性或失活状态,这表明 MCI 2 III 2 IV 2从活性状态(状态 1)转变为失活状态(状态 1)。 2),通过中间体(Mid 1、Mid 2 和 Mid 3)。
状态 1 中 2Fe-2S 团簇的动态范围与“b”状态中报告的动态范围基本一致; Mid 1 的动态范围与“int”状态的动态范围基本一致;状态 2 的动态范围在“int”和“c1”状态之间重叠。因此,当 MCI 2 III 2 IV 2从状态 1 转变为状态 2 时,CIII 的 Rieske 域从“b”状态转变为“c1”状态。
此外,在状态 1 和中 1 中,CIV 的 Cyt.c 结合位点与 CIII 的两个 Cyt.c 结合位点之间的距离分别约为 95 Å 和 115 Å,而在状态 2 中,这些距离约为分别为 73 埃和 78 埃。这一发现表明,Cyt.c 可能在状态 2 中 Cyt.c 结合位点之间的距离明显更短的情况下更容易扩散。此外,我们注意到 Cyt.c 的密度逐渐从状态 1 分散到中间 1,再到状态 1。状态 2,表明 Cyt.c 在状态 1 中与 CIII 结合更紧密,而在状态 2 中结合更松散。
随着 MCI 2 III 2 IV 2从状态 1 转变为状态 2,IMS 侧 CI 和 CIII 之间的两个相互作用点发生显着的构象变化。特别地,NDUFB7的长螺旋(C80-G126)从CI膜臂的远端延伸以接触CIII单体A的Rieske结构域,并且NDUFB10的C末端从CI膜臂的中部区域突出接触 CIII 单体 B 的 UQCRH N 端区域。在状态 1 中,这两个相互作用位点之间的最近距离分别为 ~15.3 Å 和 ~11.9 Å,而在状态 2 中,这些距离减小到 ~6.0 Å和~7.9 Å,分别。 Rieske 结构域对于 CIII 内的电子转移至关重要,UQCRH 的 N 端区域与 CIII 的 Cyt.c 结合位点相邻。因此,CI 的 NDUFB7 A123和 NDUFB10 A176可能充当两个锚定点,诱导 CIII Rieske 结构域和 Cyt.c 结合位点的构象变化。